10.31047/1668.298x.v1.n40.35653
Articulos
Evaluación de variables agronómicas en el cultivo de vara de oro (Solidago chilensis Meyen) en la pampa húmeda argentina
B. Perez
B. A. Lus
G. N. Barrientos
D. M. Carrettoni
N. M. Apóstolo
1 Perez, B., (ORCID: 0000-0002-2240-3631), Lus, B. A. (ORCID: 00000002-6434-3446), y Apóstolo, N. M. (ORCID: 0000-0002-3763-3915): Universidad Nacional de Luján (UNLu), Departamento de Ciencias Básicas, Instituto de Ecología y Desarrollo Sustentable (INEDES-UNLu-CONICET), Programa Interdisciplinario de Estudios en Plantas Vasculares (PIEPVAS). Luján, Buenos Aires, Argentina. Barrientos, G. N. (ORCID: 0000-0002-5291-669X), Universidad Nacional de Luján (UNLu), Departamento de Ciencias Básicas, Instituto de Ecología y Desarrollo Sustentable (INEDES-UNLu-CONICET), Programa de Investigación en Ecología Terrestre (PIET). Luján, Buenos Aires, Argentina. Carrettoni, D. M. (ORCID: 0000-0002-4130-2414), Universidad Nacional de Luján. Luján, Buenos Aires, Argentina. Correspondencia a: perezbea99@gmail.com
RESUMEN
Vara de oro (Solidago chilensis) es una especie herbácea perenne nativa de Argentina. Aunque presenta importantes propiedades medicinales y potencial valor ornamental, esta especie no tiene producción en la pampa húmeda. El presente trabajo evalúa el efecto de dos densidades de siembra (71 500 y 143 000 plantas.ha-1, campaña 2013-2014) y dos fechas de siembra (temprana en septiembre y tardía en noviembre, campaña 2014-2015) en la fenología, caracteres morfométricos y rendimiento de materia seca de dos cosechas por ciclo de cultivo. Se determinó que la fenología y los rendimientos de ambas cosechas no fueron afectados por la densidad de siembra. Sin embargo, la menor densidad disminuyó la longitud del tallo principal y aumentó el largo de la inflorescencia principal y el número de ramas basales. La siembra temprana alargó el período vegetativo y el reproductivo, e incrementó la longitud del tallo principal, número de inflorescencias, número de ramas basales y rendimientos de la primera y segunda cosecha. En ambas campañas, la primera cosecha produjo mayores rendimientos que la segunda. Como prácticas de manejo para el cultivo de vara de oro en la región, se recomienda la siembra en septiembre con una densidad de 71 500 plantas.ha-1 y una sola cosecha por ciclo.
Palabras clave: caracteres morfométricos, fenología, floricultura, plantas medicinales, rendimiento.
SUMMARY
Goldenrod (Solidago chilensis) is a perennial herbaceous species native to Argentina. Even though it has important medicinal properties and potential ornamental value, the species is not produced in the Humid Pampas. We evaluated the effect of two sowing densities (71 500 and 143 000 plants.ha-1, during 2013-2014 season) and two sowing dates (early sowing in September and late sowing in November, during 2014-2015 season) on phenology, morphometric characters and dry matter yield of two harvests per cultivation cycle. Plant density did not affect phenology or yields of both harvests, but lower plant density reduced main stem length and increased main ¡nflorescence length and number of basal branches. Early sowing lengthened the vegetative and reproductive growth stages and increased main stem length, number of ¡nflorescences, number of basal branches and yields of the first and second harvest. In both seasons, the first harvest produced higher yields. Sowing in September with a density of 71 500 plants.ha-1 and a single harvest per cycle are recommended management practices for cultivation of goldenrod in the region.
Keywords: morphometric characters, phenology, floriculture, medicinal plants, yield.
INTRODUCCION
El género Solidago L. (Asteraceae) incluye, aproximadamente, 139 especies de plantas herbáceas perennes conocidas como vara de oro. En su mayoría, están distribuidas en América del Norte y, escasamente, en Sudamérica, Europa y Asia (Lopez Laphitz y Semple, 2015; Semple, 2022). Los miembros del género son reconocidos por sus propiedades medicinales y su valor ornamental. Sin embargo, las producciones comerciales no se encuentran extendidas ampliamente. En la medicina tradicional europea, se destaca Solidago virgaurea L., una especie originaria de Eurasia y, actualmente, presente en todo el mundo. En determinados países se cultiva comercialmente para su utilización como droga cruda, pero a escala limitada (Kotodziej, 2009). Por este motivo, se realizaron numerosos estudios agronómicos para obtener rendimientos locales (Galambosi et al., 1993; Muñoz Centeno, 2003; Gruszczyk y
Las especies de Solidago exhiben llamativas inflorescencias amarillas con excelentes cualidades para su uso como complemento floral y macizos florales. En los últimos años, países productores de flores de corte, como Holanda e Israel, desarrollaron numerosos cultivares, principalmente de Solidago canadensis L., además de híbridos intragenéricos e intergenéricos (Romero González y García Re, 2001). Algunos de ellos fueron introducidos para el desarrollo de cultivos comerciales en países donde la especie no se considera originaria, razón por la cual se requirieron estudios para su adaptación en condiciones agroclimatológicas particulares (Romero González y García Re, 2001; Cuesta et al., 2006; Mayalica Dalgo, 2014; Yumbla Orbes et al., 2017; Collaguazo Lita, 2019).
En Argentina, habitan tres especies nativas de Solidago. S. chilensis Meyen, popularmente denominada vara de oro, vara dorada, romerillo amarillo, penacho y punta de flecha, es la que presenta mayor distribución. Abarca gran parte del territorio argentino y países limítrofes (Lopez Laphitz y Semple, 2015; Zuloaga et al., 2018). La planta tiene una altura entre 60 y 250 cm (Fernández et al., 2016), es rizomatosa, con tallos erectos simples y una inflorescencia cimosa apanojada piramidal que porta numerosos capítulos radiados. Crece en ambientes ruderales, tanto en suelos ricos como poco fértiles. Vegeta desde mediados o fines de primavera y verano y florece desde primavera hasta otoño (Fernández et al., 2016).
Las partes aéreas, tanto vegetativas como reproductivas, y las subterráneas de S. chilensis son utilizadas en medicina popular en Sudamérica para el tratamiento de diversas afecciones. Es empleada como medicamento antiinflamatorio, vulnerario, diurético y antilítico, entre otros usos (Lahitte et al., 1998; Mercandeli et al., 2012; Alonso y Desmarchelier, 2015). Por otra parte, se ha demostrado que la especie posee actividad biológica gastroprotectora, antiinflamatoria, antibacterial, antifúngica, antioxidante, antirreumática y para el tratamiento de lumbago (Güntner et al., 1999; Vila et al., 2002; Barboza et al., 2009; Tamura et al., 2009; Bucciarelli et al., 2010; da Silva et al., 2010; Mercandeli et al., 2012; Alonso y Desmarchelier, 2015). En Brasil, es considerada de interés farmacéutico por el Ministerio de Salud y se cultiva experimentalmente por la Plataforma Agroecológica de Fitomedicamentos (Valverde et al., 2012). No obstante, la mayor parte de los productos naturales para uso etnomédico que se comercializan en mercados locales se obtiene por recolección directa (Basualdo et al., 2004), con la consecuente falta de homogeneidad en los productos cosechados y la pérdida de poblaciones silvestres.
S. chilensis posee atractivas y fragantes inflorescencias que le confieren gran potencial decorativo. Ofrecen una alternativa para diversificar las producciones locales en el sector de la floricultura en Argentina, donde el uso de especies nativas ornamentales se encuentra aprovechado escasamente. La especie presenta aptitud para flor de corte y flor seca, tanto por el aspecto de las varas como por la buena conservación en florero (Cuesta et al., 2006). Además, por sus características morfológicas y anatómicas xeromórficas resulta apropiada para el paisajismo en zonas áridas y secas, donde puede mantenerse con mínimos cuidados (Gil et al., 2012; Cuesta et al., 2021). En este sentido, Imhof et al. (2011) caracterizaron varios ecotipos provenientes de la provincia de Córdoba según un índice de aptitud de uso ornamental basado en aspectos morfológicos de las inflorescencias.
Los antecedentes relacionados con el cultivo de S. chilensis incluyen, principalmente, estudios de germinación de semillas y de propagación. La especie tiene abundante producción de frutos de dispersión anemócora, aunque los porcentajes de germinación de sus semillas son variables y, por lo general, no mayores al 20 % (Correia et al., 1999). Sin embargo, los tratamientos pregerminativos de imbibición o lixiviación en agua mejoran estos valores de germinación y logran máximos entre 56 y 66,5 % (Lobler, 2013; Lus et al., 2013; Lobler et al., 2016).
En Brasil, se realizaron diversos trabajos de multiplicación de S. chilensis a partir de estacas de tallos aéreos y de rizomas, en condiciones de laboratorio, macetas y bajo invernáculo (Lobler, 2013; Lobler et al., 2013). En Argentina, Gil et al. (2017) obtuvieron plántulas a partir de estacas en ensayos realizados en invernáculo, que luego trasplantaron a jardines de bajo mantenimiento en la provincia de Córdoba. Las técnicas de propagación vegetativa resultan más apropiadas cuando las especies tienen baja producción de semillas, se presentan dificultades con la germinación o tienen baja tasa de supervivencia de plántulas en el campo; o bien, se prefiere mayor uniformidad en las plantas producidas. Por otro lado, Cuesta et al. (2006) obtuvieron, exitosamente, plántulas de la especie a partir de semillas cultivadas en macetas.
Para aprovechar el valor medicinal u ornamental de S. chilensis en la pampa húmeda, resulta indispensable estudiar su comportamiento en condiciones particulares de cultivo con el fin de definir el manejo adecuado que permita obtener una mejor calidad y homogeneidad en el material cosechado. Las referencias de prácticas culturales de la especie a campo en la región son escasas (Carrettoni, 2016; Barrientos, 2017). En este marco, los objetivos del presente trabajo fueron: a) evaluar el efecto de dos densidades (primera campaña) y dos fechas de siembra (segunda campaña) en la fenología, caracteres morfométricos y rendimiento de materia seca de dos cosechas por ciclo de cultivo de S. chilensis en la pampa húmeda; y b) realizar un registro preliminar de las enfermedades y plagas asociadas a este cultivo.
MATERIALES Y MÉTODOS
Cultivo y cosecha
Los ensayos se real izaron durante dos campañas, 2013-2014 y 2014-2015, en el campo experimental de la Universidad Nacional de Luján (34° 35' S, 59° 04' O, 28 m snm), ubicado en Luján, provincia de Buenos Aires, en la pampa húmeda de la región pampeana. El suelo de las parcelas experimentales presentaba las siguientes características: Argiudol típico, familia fina, illítica, térmica, conformado por una asociación de las Series Mercedes (50 %), Portela (40 %) y Gowland (10 %) (Instituto Nacional de Tecnología Agropecuaria, 2015). El clima de la zona es templado húmedo, con una precipitación media anual de 1050 mm, de mayor incidencia en verano. La temperatura media anual es de 16,5 °C, la media estival de 23 °C y la invernal de 5 °C (Carballo y Goldberg, 2014).
En el ensayo de la primera campaña (20132014), se analizaron dos densidades de siembra: 71 500 plantas.ha-1 (D1), con 70 cm entre líneas de siembra, y 143 000 plantas.ha-1 (D2), con 35 cm entre líneas de siembra; ambas densidades con 20 cm entre plantas. La siembra se realizó a fines de invierno (2-9-2013). En el ensayo de la segunda campaña (2014-2015), se evaluaron dos fechas de siembra: siembra temprana a fines de invierno (S1), realizada el 3-9-2014; y siembra tardía a fines de primavera (S2), realizada el 27-11-2014; ambas con 71500 plantas.ha-1, 70 cm entre líneas de siembra y 20 cm entre plantas. Las distancias entre líneas y plantas son las que se utilizan convencionalmente en diversos cultivos.
El diseño experimental se realizó en bloques completos al azar con cuatro repeticiones. La superficie de cada parcela fue de 5,6 m2 (2,8 m x 2,0 m). El terreno se preparó con varias pasadas de rastra de discos y refinado con motocultivador. Los surcos se trazaron con azada. La siembra fue manual y directa en líneas continuas a muy escasa profundidad, debido a que las semillas de la especie presentan respuesta fotoblástica positiva (Correia et al., 1999). Una vez establecidas las plántulas, se ralearon para lograr las densidades deseadas. Se efectuó desmalezamiento manual durante todo el ciclo del cultivo y se aplicó riego complementario, según las condiciones de humedad del suelo, sin agregado de fertilizantes.
En el primer ensayo, se utilizó un pool de semillas provenientes de poblaciones silvestres de S. chilensis de los alrededores del campo experimental. En el segundo ensayo, se emplearon semillas cosechadas del primer ensayo. Las semillas fueron almacenadas en bolsas de papel a 4 °C hasta el inicio de los ensayos. Antes de la siembra, fueron sometidas a un tratamiento de lixiviación durante 24 h con agua corriente en circulación continua.
Con el fin de maximizar el rendimiento del cultivo, se efectuaron dos cosechas por ciclo y se llevaron a cabo en la fase de plena floración en las filas centrales de las parcelas. La primera cosecha tuvo modalidades distintas, según el ensayo. En el primer ensayo, se cortaron las partes aéreas de las plantas (tallos, hojas e inflorescencias) a 10 cm sobre el suelo, para favorecer el rebrote. En el segundo ensayo, se cortaron las inflorescencias únicamente, dejando en pie la parte vegetativa. Luego de la cosecha, se cortó el resto de las plantas de cada parcela para que todos los individuos quedaran en igualdad de condiciones. En la segunda cosecha, se extrajeron plantas completas. Las fechas de la primera y segunda cosecha para el primer ensayo fueron: 6-3-2014 y 4-8-2014, respectivamente. Para el segundo ensayo, las fechas de la primera y segunda cosecha fueron, respectivamente: 13-3-2015 y 4-5-2015 (para la siembra temprana), y 11-5-2015 y 24-6-2015 (para la siembra tardía). El material cosechado se separó en distintas fracciones, según la cosecha: fracción reproductiva (inflorescencias), fracción vegetativa aérea (tallos y hojas) y fracción vegetativa subterránea (raíces y rizomas). El corte de inflorescencias se efectuó a 5 cm por debajo de la ramificación basal. Para la extracción de raíces y rizomas, se removió el suelo por completo en los primeros 20 cm de profundidad.
Medición y análisis de variables
Las observaciones fenológicas se realizaron una vez por semana sobre 10 plantas seleccionadas al azar de las filas centrales de las parcelas. Con la escala fenológica BBCH (Enz y Dachler, 1997), se identificaron las siguientes fases: emergencia (aparición de cotiledones a través de la superficie del suelo), prefloración (botones florales visibles), 10 % de floración o inicio de floración (10 % de capítulos con flores abiertas), 40 % de floración (40 % de capítulos con flores abiertas) y 50 % de floración o plena floración (50 % de capítulos con flores abiertas). Se registró el inicio de las fases en días después de la siembra y se consideraron alcanzadas cuando el 50 % de las plantas presentaban las características de la fase indicada. Se consideró el período vegetativo desde la fase de emergencia hasta el inicio de prefloración.
Las mediciones morfométricas se efectuaron en fase de plena floración al momento del primer corte en cinco plantas de los surcos centrales seleccionadas aleatoriamente. Los caracteres evaluados fueron: longitud del tallo principal (medido desde el cuello hasta 5 cm por debajo de la ramificación basal de la inflorescencia principal o terminal), número de ramas laterales (ramificaciones mayores a 10 cm de longitud) formadas a lo largo del tallo principal, número de ramas basales (ramificaciones laterales formadas cerca del cuello de la planta), número de inflorescencias (incluyendo la principal y las originadas en ramas laterales y basales), longitud de la inflorescencia principal (medida sobre su eje primario desde el ápice hasta 5 cm por debajo la ramificación basal) y número de ramificaciones (mayores a 5 cm de longitud) de la inflorescencia principal. Antes de la segunda cosecha, se realizaron observaciones morfológicas cualitativas de las inflorescencias laterales, ya que las plantas no produjeron nuevas inflorescencias principales luego del primer corte.
Para el cálculo del rendimiento de materia seca, en primer lugar, se registró el peso fresco de las muestras de las fracciones cosechadas en cada parcela. Luego, se tomaron submuestras de 50-80 g, que fueron colocadas en estufa a 60 °C hasta peso constante para obtener el peso seco. Finalmente, estos datos se extrapolaron a rendimientos por superficie (kg.ha-1) para cada tratamiento.
Con el objetivo de realizar una primera caracterización sanitaria del cultivo, se identificaron las enfermedades y plagas. Durante las diferentes fases fenológicas, se realizaron observaciones semanales de los tallos, hojas e inflorescencias (raíces y rizomas se analizaron al momento de la cosecha) de 10 plantas de cada tratamiento, para registrar la presencia de insectos y de daños visibles generados por insectos o patógenos. Se tomaron muestras de partes vegetales afectadas y se recolectaron los insectos para su identificación (Laboratorio de Fitopatología y Zoología Agrícola de la Universidad Nacional de Luján).
Los datos de morfometría (tomados antes de la primera cosecha) y de rendimiento de materia seca (tomados en dos cosechas) fueron procesados mediante análisis de varianza (p < 0,05) con el paquete estadístico SPSS 15.0. Los efectos de la densidad de siembra (dos tratamientos, primera campaña) se analizaron independientemente de la fecha de siembra (dos tratamientos, segunda campaña). En cada campaña, los rendimientos de cada cosecha se evaluaron por separado.
RESULTADOS Y DISCUSIÓN
Fenología
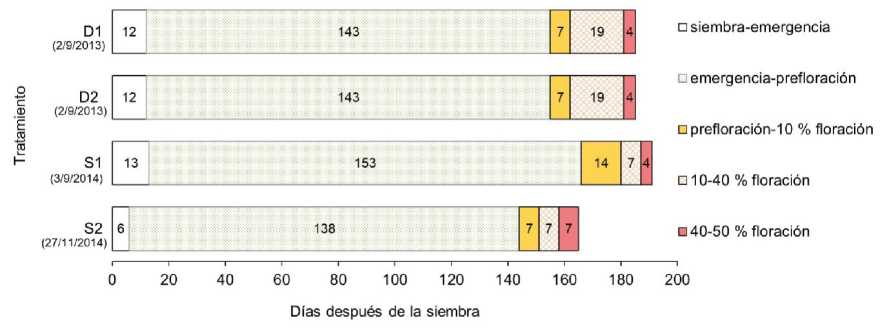
Las fases fenológicas no mostraron diferencias entre los cultivos de menor y mayor densidad de siembra (Figura 1). La emergencia ocurrió 12 días después de la siembra y el período vegetativo se extendió por 155 días. Los cultivos iniciaron la fase de floración una semana después del comienzo de la prefloración (Figura 2B), a los 162 días de la siembra, y llegaron a fase de plena floración en 23 días. La duración del período reproductivo fue de 30 días y la del ciclo de cultivo fue de 185 días. El rebrote desarrollado durante los meses de otoño e invierno a partir de una porción basal del tallo principal no presentó buen crecimiento vegetativo. Además, pocas plantas alcanzaron el período reproductivo durante los cinco meses previos a la segunda cosecha. En condiciones diferentes a las de este ensayo (en producciones bajo invernáculo, con aplicación de fertilizantes, temperatura y fotoperíodo controlados, pinzado y poda), los tallos de S. canadensis rebrotados a partir de rizomas pueden rendir hasta cuatro cortes de varas florales por ciclo de cultivo (Romero González et al., 2001).
Figura 1: Duración de las fases fenológicas en cultivos de Soiidago chiiensis bajo dos densidades de siembra (ensayo campaña 20132014) y dos fechas de siembra (ensayo campaña 2014-2015) en Luján, Buenos Aires
En el cultivo temprano del segundo ensayo, las fases fenológicas fueron de mayor o igual duración que las del cultivo tardío (Figura 1), con excepción de la fase 40-50 % de floración (Figura 2C), que fue más breve. Para la emergencia, se registró un incremento de 7 días con respecto al cultivo tardío. La fase emergencia-prefloración (Figura 2A) fue 15 días más extensa, y el inicio de la floración ocurrió en el doble de tiempo. El período reproductivo duró 25 días en siembra temprana y 21 días en siembra tardía. El ciclo del cultivo temprano se extendió por 191 días, desde fines de invierno hasta fines de verano. Por otro lado, el cultivo tardío se desarrolló desde fines de primavera hasta mediados de otoño y su ciclo fue 25 días más corto que el del cultivo temprano (Figura 1). Luego del corte de inflorescencias, se obtuvo una segunda cosecha de tallos floríferos en plena floración (Figura 2F, G) en el término de 52 días en el cultivo temprano y 44 días en el tardío. Con esta modalidad de cosecha se lograron dos floraciones por ciclo de cultivo a campo en la pampa húmeda.

Figura 2: Parcelas experimentales de Soiidago chiiensis en distintas fases fenológicas y morfología de las plantas cultivadas en ensayos realizados en Luján, Buenos Aires. A: Cultivo de siembra temprana (detrás) y tardía (adelante) en estado vegetativo (emergencia-prefloración). B: Cultivo de mayor densidad de siembra en prefloración. C: Cultivo de siembra temprana en plena floración. D: Sector del vástago de un ejemplar del cultivo temprano con ramas basales. E: Planta entera con inflorescencia principal y laterales (cultivo de siembra temprana en primera plena floración). F: Cultivo de siembra temprana en segunda plena floración. G: Tallos principales con inflorescencias laterales de la segunda floración (cultivo de siembra temprana)
Aunque los cultivos del primer ensayo (20132014) y el de siembra temprana del segundo ensayo (2014-2015) se iniciaron en el mismo momento del año, presentaron diferencias en la fenología evaluada. El ciclo de los cultivos del primer ensayo fue ligeramente inferior al del cultivo del segundo ensayo; el período vegetativo fue más corto y el período reproductivo fue más largo. Además, en el primer ensayo, las fases emergencia-prefloración y prefloración-inicio de floración fueron de menor duración que en el segundo ensayo, y la fase 1040 % de floración (Figura 1) fue más extensa. La variabilidad en las condiciones meteorológicas entre los años de los ensayos o el diferente origen de las semillas empleadas en ambos casos podrían explicar las diferencias observadas en la fenología.
Caracteres morfométricos
La densidad de siembra afectó significativamente la longitud del tallo principal, la longitud de la inflorescencia principal y el número de ramas basales (Tabla 1). A menor densidad, las plantas produjeron un tallo principal más corto, inflorescencias 41,4 % más largas y, aproximadamente, el doble de ramas basales que las plantas en mayor densidad de siembra. Estos resultados concuerdan con los obtenidos en ensayos a campo con S. virgaurea bajo distintos distanciamientos entre líneas de siembra (30, 40 y 50 cm), en los que la producción de ramas por planta se incrementaba con el aumento del espaciamiento entre líneas de plantas (Kotodziej, 2008a). La respuesta de las plantas a las bajas densidades resulta, frecuentemente, en aumentos en los valores de determinadas variables morfométricas vegetativas y/o reproductivas (Gil et al., 1999).
La fecha de siembra produjo diferencias significativas en la longitud del tallo principal, en el número de ramas basales y en el número de inflorescencias (Tabla 1). El tallo principal en el cultivo temprano duplicó su longitud con respecto al del cultivo tardío y produjo el doble de ramas basales y de inflorescencias (Figura 2D, E). Esto podría atribuirse al período de crecimiento vegetativo más prolongado del cultivo temprano y al aumento progresivo del fotoperíodo durante su ciclo. Según Flórez Roncancio y Pereira (1998), los días largos promueven la elongación del tallo y la inducción floral en S. chilensis, así como el aumento del número de ramas laterales y de hojas. En el ensayo en Luján, tanto el cultivo temprano como el tardío tuvieron las condiciones necesarias para la inducción floral y, una vez alcanzadas, produjeron inflorescencias principales similares, con igual longitud y número de ramificaciones, por lo que estas características no fueron afectadas por la fecha de siembra (Tabla 1).
Las inflorescencias laterales producidas en la segunda floración del segundo ensayo presentaron una arquitectura más sencilla que las de la primera floración: un eje principal muy corto con ramificaciones escasas y poco desarrolladas (Figura 2G). Incluso, se observaron inflorescencias no ramificadas. La baja tasa de crecimiento de los cultivos y la disminución del fotoperíodo luego de la primera cosecha explicarían la estructura más simple de las inflorescencias obtenidas en la segunda cosecha. En cultivos de S. canadensis para complemento floral de corte realizados con control de fotoperíodo y temperatura, las plantas rebrotadas mantienen el aspecto general de las inflorescencias, aunque la calidad de las varas disminuye en cultivos más viejos (Romero González y García Re, 2001).
Promediando las dos campañas, las plantas de S. chilensis, bajo las condiciones de cultivo ensayadas en Luján, desarrollaron un tallo principal de 61,5 cm de largo hasta la inflorescencia principal y de 51,3 cm en la zona florífera terminal. A su vez, los tallos produjeron 3,7 ramas basales y 8,7 ramas laterales, 9,4 inflorescencias y casi 37 ramas en la inflorescencia principal. Al comparar estas características morfométricas con las de plantas silvestres de las cercanías del presente estudio, se observan diferencias notables en la arquitectura de las plantas. Las plantas silvestres producen, en general, solo la inflorescencia principal (Barrientos, 2017), la cual es más corta y menos ramificada que la que exhiben las plantas cultivadas. Además, rara vez desarrollan ramas basales y laterales, predominando la forma de vara erecta (Barrientos, 2017). De igual manera, en un ensayo de cultivo de S. chilensis en macetas con fertilizantes, Cuesta et al. (2006) obtuvieron plantas de mayor altura, con mayor número de ramas, inflorescencias más largas y de mejor aspecto ornamental que aquellas que crecían en su hábitat natural en la provincia de Mendoza. En otro estudio similar realizado con S. virgaurea en Finlandia, Galambosi et al. (1993) encontraron que las plantas bajo cultivo fertilizado eran de mayor altura que las plantas silvestres y producían inflorescencias más largas y más ramificadas. Las prácticas culturales disminuyen la competencia intraespecífica e interespecífica al crear condiciones más favorables para el crecimiento de las plantas que aquellas que se presentan en el hábitat natural, modificando, a menudo, sus características morfológicas y fisiológicas con relación a las de las plantas silvestres (Díaz Guillén, 2010).

Tabla 1: Caracteres morfométricos (media ± desvío estándar) de Solidago chilensis bajo dos densidades de siembra (ensayo campaña 2013-2014) y dos fechas de siembra (ensayo campaña 2014-2015) en Luján, Buenos Aires
Rendimiento
En la primera cosecha del primer ensayo, no se registraron diferencias estadísticamente significativas entre las dos densidades de siembra evaluadas con respecto al rendimiento de materia seca reproductiva y vegetativa aérea (Tabla 2). El promedio de materia seca entre las dos densidades fue 1681,6 kg.ha-1 para la fracción reproductiva y 3834,5 kg.ha-1 para la fracción vegetativa. La menor densidad de plantas pudo haber sido compensada con mayor emisión de ramas basales por planta y mayor longitud de la inflorescencia principal, o bien, por otras variables no evaluadas en el presente trabajo, como el número de hojas o diámetro de tallos. De esta manera, se conseguiría una producción similar al tratamiento con mayor densidad de siembra. En contraste, la menor densidad de siembra ensayada en cultivos de S. canadensis produjo un aumento en los valores de todos los parámetros morfométricos analizados (entre ellos, altura y circunferencia del tallo vegetativo, número de hojas, y longitud y número de ramas de la inflorescencia principal), lo que dio como resultado una mayor producción de materia fresca y seca (Osman, 2014). A su vez, Kotodziej (2008a) encontró que el espaciamiento entre líneas de 30 y 40 cm en cultivos de S. virgaurea producía rendimientos de materia seca aérea superiores al distanciamiento de 50 cm.
Los rendimientos de materia seca vegetativa aérea y reproductiva de S. chilensis del primer ensayo variaron, según la densidad, entre 5025 y 6007 kg.ha-1. Estos valores son comparables a los obtenidos en cultivos de S. virgaurea iniciados a partir de semillas, en los que la cosecha de partes aéreas produjo entre 5000 y 9900 kg.ha-1 de materia seca, dependiendo de factores como el espaciamiento entre líneas de siembra, época de siembra, fertilizantes aplicados y tipo de suelo, entre otros (Galambosi et al.,1993; Kotodziej, 2008a, 2008b, 2009).
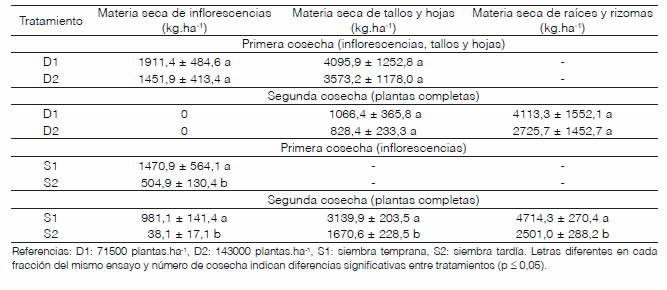
Tabla 2: Rendimiento de materia seca (media ± desvío estándar) de las fracciones reproductiva (inflorescencias), vegetativa aérea (tallos y hojas) y vegetativa subterránea (raíces y rizomas) de dos cosechas por ciclo de cultivo de Solidago chilensis bajo dos densidades de siembra (ensayo campaña 2013-2014) y dos fechas de siembra (ensayo campaña 2014-2015) en Luján, Buenos Aires
En la segunda cosecha del primer ensayo, la densidad de siembra no afectó los rendimientos de materia seca aérea (Tabla 2). Sin embargo, fueron mucho más bajos que los de la primera cosecha, debido a la baja tasa de crecimiento del rebrote en otoño e invierno. La producción promedio de materia seca vegetativa aérea representó el 24,7 % de lo obtenido en la primera cosecha, mientras que la materia seca reproductiva fue considerada nula, dado que solo algunas plantas del ensayo produjeron nuevas inflorescencias. Por otro lado, la materia seca subterránea no fue afectada por la densidad de siembra (Tabla 2), con un promedio de 3419,5 kg.ha-1. El rendimiento total de las dos cosechas de este ensayo fue 11187,0 kg.ha-1 para la menor densidad y 8579,2 kg.ha-1 para la mayor densidad de siembra.
Los rendimientos de las dos cosechas del cultivo temprano del segundo ensayo fueron significativamente superiores a los del cultivo tardío (Tabla 2). En la primera cosecha, la materia seca reproductiva del cultivo temprano fue, aproximadamente, tres veces mayor que la del cultivo tardío; mientras que, en la segunda cosecha, lo superó cerca de 26 veces. No obstante, los rendimientos de la segunda cosecha fueron menores que los de la primera. La arquitectura más sencilla de las inflorescencias de la segunda floración se reflejó en menor producción de materia seca con respecto a la primera floración: 66,7 % para siembra temprana y 7,5 % para siembra tardía. Por otra parte, los rendimientos vegetativo aéreo y subterráneo del cultivo temprano casi duplicaron a los del tardío (Tabla 2). La siembra temprana permitió un crecimiento más prolongado del cultivo, aumentando la longitud del tallo principal y la cantidad de ramas basales y de inflorescencias, lo que produjo rendimientos mayores. El rendimiento total de ambas cosechas del cultivo temprano de este ensayo fue 10306,2 kg.ha-1, el cual se encuentra dentro del mismo rango de valores que los del primer ensayo, que también fueron iniciados en siembra temprana. El total de 4714,6 kg.ha-1 obtenido para el cultivo tardío representa un rendimiento mucho menor que el de los cultivos tempranos. De manera similar, en experimentos a campo en Polonia, los cultivos de S. virgaurea sembrados en otoño (siembra temprana) produjeron mayores rendimientos de materia seca que los sembrados en primavera (Kotodziej, 2008a).
Enfermedades y plagas
Una de las enfermedades detectadas fue el moho blanco, causado por Sclerotinia sclerotiorum (Lib.) de Bary. Este hongo produjo marchitamiento, necrosis y posterior muerte de algunas plantas durante el período vegetativo. Se encontraron esclerocios en los tallos afectados y se observó podredumbre húmeda causada por agentes patógenos oportunistas. Esta es una de las adversidades más frecuentes en cultivos de Solidago, causante de podredumbre de tallos y raíces (Mayalica Dalgo, 2014). Además, se hallaron hojas afectadas con roya (Puccinia sp.).
Con respecto a insectos, se encontraron especies que, habitualmente, se consideran plagas de cultivos. En el período vegetativo se hallaron adultos de curculiónidos (Coleoptera: Curculionidae), larvas de Rachiplusia nu Guenée (Lepidoptera: Noctuidae) y larvas de Mythimna adultera Schaus (Lepidoptera: Noctuidae). En las inflorescencias se hallaron adultos de Astylus atromaculatus Blanch (Coleoptera: Melyridae). Durante la segunda cosecha, se encontraron grandes cantidades de secreción blanquecina serosa de tipo filamentosa que rodeaban las raíces y rizomas de algunas plantas, característica del pulgón lanígero Eriosoma lanigerum Hausmann (Hemiptera: Aphididae). Sin embargo, no se hallaron tumores o agallas en los órganos subterráneos causados por este insecto.
CONCLUSIONES
El cultivo de S. chilensis se realizó exitosamente en la pampa húmeda a partir de siembra directa de semillas. Se determinó que las etapas fenológicas del cultivo fueron afectadas por la fecha de siembra. Además, algunas de las variables morfométricas analizadas fueron modificadas por la densidad de siembra, fecha de siembra, o ambas. Los rendimientos de materia seca de plantas cosechadas en plena floración no fueron afectados por la densidad de siembra. Sin embargo, los rendimientos de la siembra temprana fueron superiores a los de la siembra tardía en las dos cosechas del ciclo de cultivo. Las segundas cosechas tuvieron menores rendimientos que las primeras y las inflorescencias obtenidas fueron de baja calidad ornamental. Por consiguiente, se recomienda la siembra en septiembre de 71500 plantas.ha-1 con 70 cm entre líneas y una sola cosecha por ciclo de cultivo para la zona de la pampa húmeda.
AGRADECIMIENTOS
Agradecemos a los especialistas de las asignaturas Fitopatología y Zoología agrícola del Departamento de Tecnología de la Universidad Nacional de Luján, por la identificación de hongos e insectos en los cultivos. Esta investigación fue financiada con el subsidio de investigación finalidad 3.5 del Departamento de Ciencias Básicas de la Universidad Nacional de Luján.
Fecha de recepción: 14/09/2022
fecha de aceptación: 03/04/2023
BIBLIOGRAFÍA
Alonso, J. y Desmarchelier, C. (2015). Plantas medicinales autóctonas de la Argentina: bases científicas para la aplicación en atención primaria de la salud (3a ed.). Corpus.
Barboza, G., Cantero, J., Nuñez, C., Pacciaroni, A. y Ariza Espinar, L. (2009). Medicinal plants: A review and a phytochemical and ethnopharmacological screening of a native Argentine Flora. Kurtziana, 34(1-2), 7-365. https://ri.conicet.gov.ar/handle/11336/24328
Barrientos, G. N. (2017). Estudios agronómicos para la domesticación de Solidago chilensis var chilensis (Asteracaee), especie nativa de interés medicinal. Trabajo final de grado. Universidad Nacional de Luján. http://ri.unlu.edu.ar/xmlui/handle/rediunlu/504 Basualdo, I., Soria, N., Ortíz, M. y Degen, R. (2004). Plantas medicinales comercializadas en los mercados de Asunción y Gran Asunción. Parte I. Rojasiana, 6(1), 95-114.
Bucciarelli, A., Minetti, A., Milczakowskyg, C. y Skliar, M. (2010). Evaluation of gastroprotective activity and acute toxicity of Solidago chilensis Meyen (Asteraceae). Pharmaceutical Biology, 48(9), 10251030. https://doi.org/10.3109/13880200903453131 Carballo, C. S. y Goldberg, S. (2014). Comunidad e información ambiental del riesgo. Las inundaciones y el río Luján. Dunken.
Carrettoni, D. M. (2016). Estudios preliminares para la domesticación de Solidago chilensis Meyen var chilensis (Asteracaee), una especie medicinal nativa. Trabajo final de grado no publicado. Universidad Nacional de Luján.
Collaguazo Lita, N. M. (2019). Determinación del efecto de los elementos climáticos sobre el crecimiento, desarrollo y producción de Solidago canadensis y Delphinium elatum. Pichincha. Tesis de grado. Universidad Central del Ecuador. http://www.dspace. uce.edu.ec/handle/25000/20181
Correia, E., Ming, L. C. y C mara, F. L. A. (1999). Aspects of the sexual reproduction of the Brazilian arnica (Solidago chilensis Meyen var. megapotamica (DC)
Cabrera-Asteraceae. Acta Horticulturae, 502, 89-92. https://doi.org/10.17660/ActaHortic.1999.502.11
Cuesta, G., Fioretti, S., Lorello, I., Pereyra, A., Gutiérrez M. T., Cabral, J. M., Pisi, G., Baglio, C. y Di Cenzi, C. (2021). Plantas nativas del centro oeste argentino con aptitud ornamental: experiencias en Mendoza. En G. Facciuto y M. Pérez de la Torre (Eds.), Plantas nativas ornamentales de Latinoamérica. Experiencias hacia la puesta en valor (73-90). INTA. http://hdl.handle. net/20.500.12123/10744
Cuesta, G., Vespa, J., Sánchez, V., Videla, E., Fioretti, S. y Cialli, A. (2006). Estudios preliminares sobre la adaptación de “penacho amarillo” (Solidago chilensis) con fines ornamentales. En Actas del III Congreso Argentino de Floricultura y VIII Jornadas Nacionales de Floricultura (s. p.). Buenos Aires, Argentina: INTA.
da Silva, A. G., de Sousa, C. P. G., Koehler, J., Fontana, J., Christo, A. G. y Guedes-Bruni, R. R. (2010). Evaluation of an extract of Brazilian arnica (Solidago chilensis Meyen, Asteraceae) in treating lumbago. Phytotherapy Research, 24, 283-287. https://doi. org/10.1002/ptr.2934
Díaz Guillén, F. (2010). El proceso de domesticación en las plantas. Casa del Tiempo, 3(28), 66-69. https://www. uam.mx/difusion/casadeltiempo/28_iv_feb_2010/ casa_del_tiempo_eIV_num28_66_70.pdf
Enz, M. y Dachler, C. H. (1997). Compendium of growth stage Identification keys for mono- and dicotyledon-ous plants. Extended BBCH scale. BBA, BSA, IGZ, IVA, AgrEvo, BASF, Bayer y Novartis.
Fernández, O. A., Leguizamón, E. S. y Acciaresi, H. A. (Eds.). (2016). Malezas e invasoras de la Argentina. Tomo II: Descripción y reconocimiento. Universidad Nacional del Sur.
Flórez Roncancio, V. J. y Pereira, M. F. A. (1998). Influencia del fotoperíodo en el desarrollo floral de plantas de Solidago chilensis, Aster ericoides cv. “Montecasino” y Solidago x luteus. Agronomía Colombiana, 15(1), 82-97. https://revistas.unal.edu.co/index.php/agrocol/ article/view/21500
Galambosi, B., Honkala, Y. y Jokela, K. (1993). Production potential of wild and cultivated golden rod (Solidago virgaurea L.) populations in Finland. Acta Horticulturae, 344, 249-255. https://doi.org/10.17660/ ActaHortic.1993.344.29
Gil, A., de la Fuente, E., Lenardis, A., Lorenzo, S. y Marengo, S. (1999). Coriander (Coriandrum sativum L.) yield response to plant population. Journal of Herbs, Spices and Medicinal Plants, 6(3), 63-73. https://doi.org/10.1300/J044v06n03_06
Gil, S. P., Reyna, M. E., Seisdedos, L. y Argüello, I. P. (2017). Reproducción de Solidago chilensis, asterácea autóctona apta para jardinería de bajo mantenimiento. Phyton, 86, 340-344. https://pdfs.semanticscholar.org/4e44/12395bcbc4cf4ec048a76e0be92a568174a2.pdf
Gil, S. P., Seisdedos, L., Reyna, M. E. y Cerana, M. M. (2012). Epidermis foliar de tres especies de asteráceas nativas de Argentina con potencial ornamental. Phyton, 81, 205-210. http://www.scielo.org.ar/scielo. php?pid = S1851-56572012000200011&script=sci_ arttext&tlng=pt
Gruszczyk, M. y Kieltyka, A. (2005). Yields and raw material quality of Hypericum perforatum L. and Solidago virgaurea L. from one-year and two-years plantation. Herba Polonica, 51, 5-9.
Güntner, C., Barra, C., Cesio, M. V., Dellacassa, E., Ferrando, L., Ferreira, F., García, C., González, G., Heinzen, H., Lloret, A., Lorenzo, D., Menéndez, P., Paz, D., Soule, S., Vázquez, A. y Moyn, P. (1999). Antioxidant properties of Solidago chilensis L. flavonoids. Acta Horticulturae, 501, 159-163. http:// doi.org/10.17660/ActaHortic.1999.501.23
Imhof, L., Suárez, M., Paganelli, F, Bettolli, F., Hick, E. y Facciuto, G. (2011). Caracterización de ecotipos de Solidago chilensis nativos de Argentina: índice de aptitud ornamental para complementos del ramo de flor. Horticultura Argentina 30(73), 146.
Instituto Nacional de Tecnología Agropecuaria. (2 de diciembre de 2015). Cartas de suelos de la provincia de Buenos Aires. Carta Nro. 3560-11-4 Luján. Ministerio de Agricultura, Ganadería y Pesca. https://inta.gob.ar/documentos/carta-de-suelos-de-la-provincia-de-buenos-aires
Kotodziej, B. (2008a). Effect of agrotechnical factors on the yield of goldenrod (Solidago virgaurea L. ssp. virgaurea). Herba Polonica, 54, 28-34. http://www. herbapolonica.pl/magazines-files/1282737-Herb%20 Pol%202008%2054(4)%20s.%2028-34.pdf
Kotodziej, B. (2008b). The effect of plantation establishment method and Atonik application in goldenrod (Solidago virgaurea L. ssp. virgaurea) cultivation. Acta Scientiarum Polonorum, Hortorum Cultus, 7(3), 33-39. https://czasopisma.up.lublin.pl/ index.php/asphc/article/view/3704
Kotodziej, B. (2009). Optimalisation of mineral fertilization in goldenrod (Solidago virgaurea L. ssp. virgaurea) cultivated for pharmaceutical purposes. Acta Scientiarum Polonorum, Hortorum Cultus, 8(1), 2736. https://czasopisma.up.lublin.pl/index.php/asphc/ article/view/3453
Lahitte, H. B., Hurrell, J. A., Belgrano, M. J., Jankowski, L. S., Haloua, M. P. y Mehltreter, K. (1998). Plantas medicinales rioplatenses. L.O.L.A.
Lobler, L. (2013). Propagagao, metabolismo secundário e genotoxicidade de Solidago chilensis Meyen (Asteraceae). Tesis de maestría. Universidade Federal de Santa María Santa María. https://repositorio.ufsm.br/handle/1/4877
Lobler, L., Rocha, B. N., Berte, R., Lucho, S. R., Fernandes, T. S., Soriani, H. y Paranhos, J. (2016). Comportamiento germinativo das sementes de Solidago chilensis Meyen (Asteraceae). Iheringia, Série Botánica, 71(1), 43-48. https://isb.emnuvens.com.br/iheringia/article/ view/486
Lobler, L., Rocha, B. N., Berte, R., Lucho, S. R., Richter, G., Zuliani, A. J. B. y Paranhos. J. T. (2013). Micropropagagáo de Solidago chilensis Meyen (Asteraceae). En D. Bisognin (Ed.), Anais do II Simposio de Melhoramento e Propagagao Vegetativa de Plantas (28-34). Universidade Federal de Santa María.
Lopez Laphitz, R. M. y Semple, J. C. (2015). A multivariate morphometric analysis of the Solidago chilensis group in South America and related taxa in North America (Asteraceae, Astereae). Annals of the Missouri Botanical Garden, 100(4), 423-441. http://dx.doi.
org/10.3417/2014023
Lus, B. A., Rodríguez Morcelle, M. I., Gabucci, L., Jara, S., Costaguta, M. y Apóstolo, N. M. (2013). Estudios preliminares de la germinación de Solidago chilensis Meyen (Asteraceae). Boletín de la Sociedad Argentina de Botánica, 48(Supl.), 190. https://botanicaargentina.org.ar/wp-content/ uploads/2017/05/SABSuplemento2013.pdf
Mayalica Dalgo, C. P. (2014). Valoración de la aplicación de cinco dosis de ácido giberélico en el rendimiento del cultivo de Solidago (Solidago sp. golden amazone). Tumbaco, Pichincha. Tesis de grado. Universidad Central del Ecuador, http://www.dspace.uce.edu.ec/ handle/25000/2489
Mercandeli, A. A., Bessa, G. P., Ronchi, S. N., Segato, T. P. S. y Silva, A. G. da. (2012). Evidence for the safe use of the extract from the Brazilian arnica, Solidago chilensis Meyen, in Primary Health Care. Chinese Medicine, 3(1), 4-8. https://doi.org/10.4236/ cm.2012.31002
Muñoz Centeno, L. M. (2003). Vara de oro (Solidago virgaurea L.) Aspectos botánicos y terapéuticos. Revista de Fitoterapia, 3(1), 35-42. https://www. fitoterapia.net/php/descargar_documento. php?id = 4746&doc_r=sn&num_volumen = 8&secc_ volumen=5953
Osman, A. R. (2014). Improving some quantitative and qualitative characteristics of Solidago canadensis “Tara” using Cycocel and planting density under drip irrigation and lighting systems. Life Science Journal, 11 (6), 110-118. http://www.lifesciencesite.com/lsj/life1106/016_23694life110614_110_118.pdf
Romero González, M. y García Re, F. T. (2001). Introducción al cultivo de Solidago. Comunidad Autónoma de la Región de Murcia, Consejería de Agricultura, Agua y Medio Ambiente.
Semple, J. C. (4 de julio de 2022). Solidago L.: Class¡ficat¡on and illustrations of goldenrods. University of Waterloo. https://uwaterloo.ca/astereae-lab/research/goldenrods/
Tamura, E. K., Jimenez, R. S., Waismam, K., Gobbo-Neto, L., Lopes, N. P., Malpezzi Marinho, E. A. L., Marinho, E. A. V. y Farsky, S. H. P. (2009). Inhibitory effects of Solidago chilensis Meyen hydroalcoholic extract on acute ¡nflammation. Journal of Ethnopharmacology 122(3), 478-485. https://doi.org/10.1016/.jep.2009.01.029
Valverde, S. S., de Oliveira, T B. y de Souza, S. P. (2012). Solidago chilensis Meyen (Asteraceae). Revista Fitos, 7(3), 131-136. https://doi.org/10.32712/2446-4775.2012.147
Vila, R., Mundina, M., Tomi, F, Furlán, R., Zacchino, S., Cassanova, J. y Cañigueral, S. (2002). Composition and antifungal activity of the essential oil of Solidago chilensis. Planta Medica, 68(2), 164-167. https://doi. org/10.1055/s-2002-20253
Yumbla Orbes, M., Stahringer, N. I. y Barbosa, J. G. (2017). Influencia de diferentes sistemas de cultivo en la calidad del tallo y las características de macronutrientes en Solidago canadensis. Siembra, (4)1,39-50. https://doi.org/10.29166/siembra.v4i1.301 Zuloaga, F. O., Belgrano, M. J. y Anton, A. M. (Eds.). (2018). Flora Argentina y del Cono Sur. Instituto de Botánica Darwinion. hflpY/buscadorfloraargentina. edu.ar/species/details/17743